Aparato de Golgi
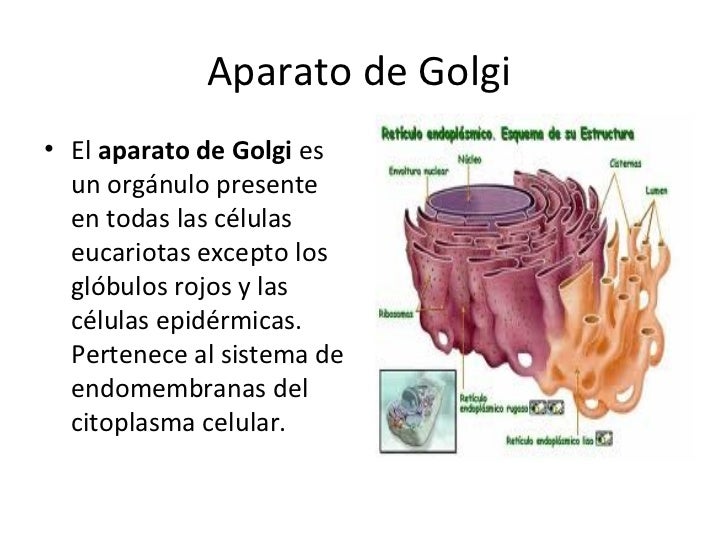
El aparato de Golgi es un orgánulo presente en todas las células eucariotas. Pertenece al sistema de endomembranas. Está formado por unos 80 dictiosomas (dependiendo del tipo de célula), y estos dictiosomas están compuestos por 40 o 60 sáculos (cisternas) aplanados y rodeados de membrana que se encuentran apilados unos encima de otros, y cuya función es completar la fabricación de algunas proteínas.
Funciona como una planta empaquetadora, modificando vesículas del retículo endoplasmático rugoso. El material nuevo de las membranas se forma en varias cisternas del aparato de Golgi.
Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos, almacenamiento y distribución de lisosomas, al igual que los peroxisomas, que son vesículas de secreción de sustancias.
La síntesis de polisacáridosde la matriz extracelular. Fue reportado por primera vez por el científico español Santiago Ramón y Cajal en el 1897, y luego descrito en gran detalle por el científico italiano Camillo Golgi, al cual debe su nombre y quien fue Premio Nobel de Medicina en 1906 junto a Santiago Ramón y Cajal.

El aparato de Golgi está compuesto por estructuras denominadas sáculos. Estas se agrupan en número variable, habitualmente de 4 a 8, formando el dictiosoma en las plantas. Presentan conexiones tubulares que permiten el paso de sustancias entre las cisternas.
Los sáculos son aplanados y curvados, con su cara convexa (externa) orientada hacia el retículo endoplasmático. Normalmente se observan entre 4 y 8, pero se han llegado a observar más de 60 dictiosomas. Alrededor de la cisterna principal se disponen las vesículas esféricas recién exocitadas.

El aparato de Golgi se puede dividir en tres regiones funcionales:
- Región Cis-Golgi: es la más interna y próxima al retículo. De él recibe las vesículas de transición, que son sáculos con proteínas que han sido sintetizadas en la membrana del retículo endoplasmático rugoso (RER), introducidas dentro de sus cavidades y transportadas por el lumen hasta la parte más externa del retículo. Estas vesículas de transición son el vehículo de dichas proteínas que serán transportadas a la cara externa del aparato de Golgi.
- Región medial: es una zona de transición.
- Región Trans-Golgi: es la que se encuentra más cerca de la membrana plasmática. De hecho, sus membranas, ambas unitarias, tienen una composición similar.
Las vesículas provenientes del retículo endoplásmico se fusionan con el cis-Golgi, atravesando todos los dictiosomas hasta el trans-Golgi, donde son empaquetadas y enviadas al lugar que les corresponda. Cada región contiene diferentes enzimas que modifican selectivamente las vesículas según donde estén destinadas. Sin embargo, aún no se han logrado determinar en detalle todas las funciones y estructuras del aparato de Golgi.
Funciones generales
La célula sintetiza un gran número de diversas macromoléculas necesarias para la vida, y el aparato de Golgi se encarga de la modificación, distribución y envío de dichas macromoléculas en la célula. Modifica proteínas y lípidos (grasas) que han sido sintetizados previamente tanto en el retículo endoplasmático rugoso como en el liso y los etiqueta para enviarlos a donde corresponda, fuera o dentro de la célula. Las principales funciones del aparato de Golgi son las siguientes:
- Modificación de sustancias sintetizadas en el RER: En el aparato de Golgi se transforman las sustancias procedentes del RER. Estas transformaciones pueden ser agregaciones de restos de carbohidratos para conseguir la estructura definitiva o para ser proteolizados y así adquirir su conformación activa. Por ejemplo, en el RER de las células acinosas del páncreas se sintetiza la proinsulina que debido a las transformaciones que sufre en el aparato de Golgi, adquirirá la forma o conformación definitiva de la insulina. Las enzimas que se encuentran en el interior de los dictiosomas son capaces de modificar las macromoléculas mediante glicosilación (adición de carbohidratos) y fosforilación (adición de fosfatos).
- Para ello, el aparato de Golgi transporta ciertas sustancias como nucleótidos y azúcares al interior del orgánulo desde el citoplasma. Las proteínas también son marcadas con secuencias señal que determinan su destino final, como por ejemplo, la manosa-6-fosfato que se añade a las proteínas destinadas a los lisosomas. Para llevar a cabo el proceso de fosforilación el aparato de Golgi importa moléculas de ATP al interior del lumen, donde las kinasas catalizan la reacción. Algunas de las moléculas fosforiladas en el aparato de Golgi son las apolipoproteínas que dan lugar a las conocidas VLDL que se encuentran en el plasma sanguíneo. Parece ser que la fosforilación de estas moléculas es necesaria para favorecer la secreción de las mismas al torrente sanguíneo.
- Secreción celular: Las sustancias atraviesan todos los sáculos del aparato de Golgi y cuando llegan a la cara trans del dictiosoma, en forma de vesículas de secreción, son transportadas a su destino fuera de la célula, atravesando la membrana citoplasmática por exocitosis. Un ejemplo de esto son los proteoglicanos que conforman la matriz extracelular de los animales. El aparato de Golgi es el orgánulo de mayor síntesis de carbohidratos. Esto incluye la producción de glicosaminoglicanos (GAGs), largos polisacáridos que son anclados a las proteínas sintetizadas en el RE para dar lugar a los proteoglicanos.
- De esto se encargarán las enzimas del Golgi por medio de un residuo de xilosa. Otra forma de marcar una proteína puede ser por medio de la sulfatación de una sulfotransferasa, que gana una molécula de azufre de un donador denominado PAPS. Este proceso tiene lugar en los GAGs de los proteoglicanos así como en los núcleos de las proteínas. Este nivel de sulfatación es muy importante para los proteoglicanos etiquetando funciones y dando una carga neta negativa al proteoglicano.
- Producción de membrana plasmática: Los gránulos de secreción cuando se unen a la membrana en la exocitosis pasan a formar parte de esta, aumentando el volumen y la superficie de la célula.
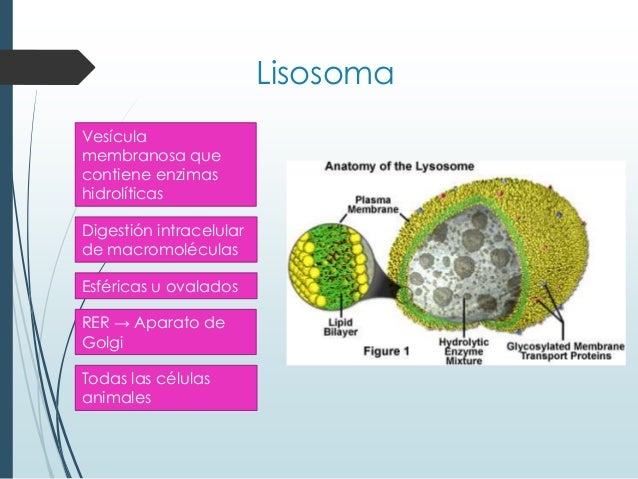
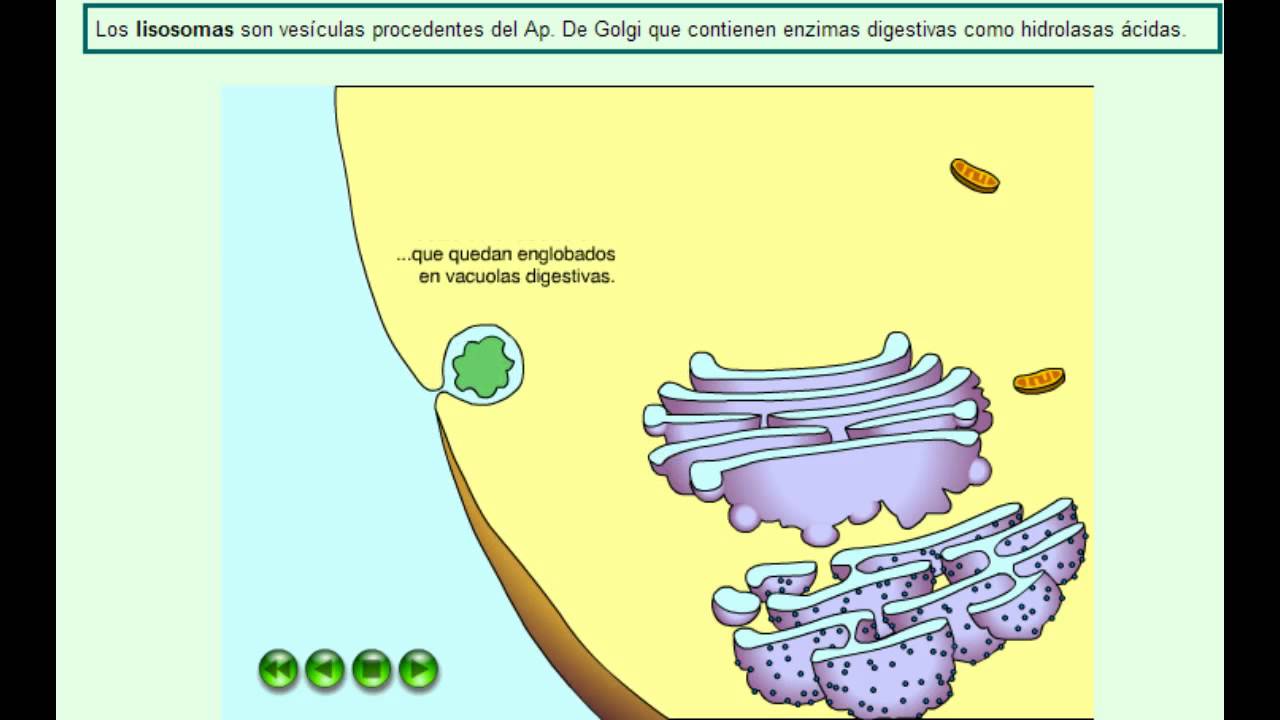
- Formación de los lisosomas primarios.
- Formación del acrosoma de los espermios.
Vesículas de transporte
Las vesículas formadas en el retículo endoplasmático liso forman, uniéndose entre ellas, agregados tubulo-vesiculares, los cuales son transportados hasta la región cis del aparato de Golgi por proteínas motoras guiadas por microtúbulos donde se fusionan con la membrana de éste, vaciando su contenido en el interior del lumen.
Una vez dentro, las moléculas son modificadas, marcadas y dirigidas hacia su destino final. El aparato de Golgi tiende a ser mayor y más numeroso en aquellas células que sintetizan y secretan continuamente sustancias, como pueden ser los linfocitos B y las células secretoras de anticuerpos.
Aquellas proteínas destinadas a zonas alejadas del aparato de Golgi son desplazadas hacia la región trans, internándose en una compleja red de membranas y vesículas asociadas denominadas región trans-Golgi.
Esta región es donde muchas proteínas son marcadas y enviadas hacia sus correspondientes destinos por medio de alguno de estos 3 tipos diferentes de vesículas, según el marcador que presenten:
| Tipo | Descripción | Ejemplo |
| Vesículas de exocitosis (constitutivas) | Este tipo de vesículas contienen proteínas que deben ser liberadas al medio extracelular. Después de internalizarse las proteínas, la vesícula se cierra y se dirige inmediatamente hacia la membrana plasmática, con la que se fusiona, liberando así su contenido al medio extracelular. Este proceso es denominado secreción constitutiva. | Los anticuerposliberados por linfocitos Bactivados. |
| Vesículas de secreción (reguladas) | Este tipo de vesículas contienen también proteínas destinadas a ser liberadas al medio extracelular. Sin embargo, en este caso, la formación de las vesículas va seguida de su almacenamiento en la célula, donde se mantendrán a la espera de su correspondiente señal para activarse. Cuando esto ocurre, se dirigen hacia la membrana plasmática y liberan su contenido como en el caso anterior. Este proceso es secreción regulada. | Liberación de neurotransmisoresdesde las neuronas. |
| Vesículas lisosomales | Este tipo de vesículas transportan proteínas destinadas a los lisosomas, unos pequeños orgánulos de degradación en cuyo interior albergan multitud de hidrolasas ácidas, lisosomas de almacenamiento. Estas proteínas pueden ser tanto enzimas digestivas como proteínas de membrana. La vesícula se fusiona con un endosoma tardío y transfiere así su contenido al lisosoma por mecanismos aún desconocidos. | Proteasasdigestivas destinadas a los lisosomas. |
Mecanismo de transporte
Los mecanismos de transporte que utilizan las proteínas para trasladarse a través del aparato de Golgi no están muy claros aún, por lo que existen diversas hipótesis para explicar dicho desplazamiento. Actualmente, existen dos modelos predominantes que no son excluyentes entre sí, hasta el punto de ser referidos a veces como el modelo combinado.

- Modelo de maduración de las cisternas: las cisternas del aparato de Golgi llevan a cabo un movimiento unidireccional desde la región cis, donde se forman, hasta la región trans, donde son destruidas. Las vesículas del retículo endoplasmático se fusionan con los dictiosomas de la región cis para dar lugar a nuevas cisternas, lo que podría generar el movimiento de las cisternas a través del aparato de Golgi a medida que se van formando nuevas cisternas en la región cis.
- Este modelo se apoya en el hecho de que se han observado al microscopio estructuras mayores que las vesículas de transporte, tales como las fibras de colágeno, desplazándose a través del aparato de Golgi. Inicialmente, esta hipótesis tuvo una gran acogida y fue la más aceptada hasta los años 80. Los últimos estudios realizados al respecto por la Universidad de Tokio y la Universidad de Chicago con tecnología más avanzada han permitido observar con mayor detalle los compartimentos y el proceso de maduración del aparato de Golgi. Además, existen evidencias de movimientos retrógrados (en dirección cis) de cierto tipo de vesículas (COP1), que transportan proteínas del retículo endoplasmático mediante el reconocimiento de péptidos señales.
- Modelo del transporte vesicular: el transporte vesicular asume que el aparato de Golgi es un orgánulo muy estable y estático, dividido en compartimentos que se disponen en dirección cis → trans. Las vesículas son las encargadas de transportar el material entre el retículo endoplasmático y el aparato de Golgi y entre los diferentes compartimentos de este.

- Las evidencias experimentales que apoyan esta hipótesis se basan en la gran abundancia de vesículas pequeñas (conocidas técnicamente como vesículas lanzadera) localizadas en las proximidades del aparato de Golgi. La direccionalidad vendría dada por las proteínas trasportadas en el interior de las vesículas, cuyo destino determinaría el movimiento de avance o de retroceso a través del aparato de Golgi, aunque también podría suceder que la direccionalidad no fuera necesaria y el destino de las proteínas viniera ya determinado desde el retículo endoplasmático. Al margen de esto, es probable que el transporte de vesículas se encuentre asociado al citoesqueleto por medio de filamentos de actina, encargados de asegurar la fusión de las vesículas con sus correspondientes compartimentos.

Las células eucariotas o eucariontes, son aquellas que poseen un núcleo definido, y se encuentran rodeados de una doble capa de lípidos, a su vez también por la envoltura nuclear, que tiene una textura porosa y resguarda el material hereditario, donde principalmente se localiza toda la información genética del ser humano.
En este tipo de células, se ubica un orgánulo llamado aparato de Golgi, el cual forma parte de un sistema de endomembranas, además se constituyo por unos 80 dictiosomas que a su vez se conforman de 40 a 60 sáculos de forma aplanada, los cuales se disponen unos encima de otros dentro de la membrana, teniendo como función la creación de varias proteínas.
¿Qué es el aparato de Golgi?
El aparato de Golgi, es aquel que funciona como una planta, donde se empacan y modifican las vesículas que constituyen el retículo endoplasmático rugoso. El material que va surgiendo de las membranas se forma en varios sáculos o cisternas del aparato de Golgi.
Este aparato se encarga de funciones como la glicosilación de las proteínas, también de los lípidos, además de su selección y destinación; y el almacenamiento y la distribución de los lisosomas y peroxisomas, los cuales son aquellas vesículas que segregan sustancias.

Estructura del aparato de Golgi
El aparato de Golgi, se conforma por las estructuras que se denominan sáculos, los cuales se agrupan variablemente de 4 a 8, que son las que hacen posible que se forme el dictiosoma en las plantas. También presentan varias conexiones tubulares, que hacen posible que pasen las sustancias entre los sáculos.
Rodeando la cisterna principal, se encuentran las vesículas que tienen forma esférica y están recién exocitadas. El aparato de Golgi consta de tres regiones eficaces:
- Región Cis Golgi, es aquella que se encuentra en un nivel mas interno y cerca del retículo, se encarga de enviar las vesículas de transición, que son aquellos sáculos de proteínas sintetizados en la membrana del retículo, además se introducen por las cavidades del mismo y se dirigen por el lumen hasta la zona externa. Estas vesículas permiten el transporte de las proteínas hacia la zona externa del aparato de Golgi.
- Región medial, es por donde pasan las vesículas de transición.
- Región Trans Golgi, está ubicada muy cerca de la membrana plasmática, teniendo una composición parecida a la misma.
Las vesículas que vienen desde el retículo endoplasmático son fusionadas a través del Cis Golgi, pasando a través de los dictiosomas hasta llegar a la región trans Golgi, allí se empaquetan y se envían hasta el lugar correspondiente.
Cada una de las regiones se conforma por diversas enzimas, que se encargan de modificar las vesículas de manera selectiva hacia su destino.
Funciones del Aparato de Golgi
- Se encarga de modificar las sustancias que son sintetizadas por el retículo endoplasmático rugoso. Esas modificaciones son agregación de restos de carbohidratos, con el fin de lograr una estructura definitiva o para que adquieran una conformación activa.
- Posibilita la secreción celular, donde las sustancias que pasan por los sáculos hasta llegar al dictiosona en forma de vesículas, son llevadas hasta su destino en el exterior de la célula, pasando por la membrana citoplasmática, debido a la exocitosis.
- Produce glicosaminoglicanos.
- Produce la membrana plasmática, cuando los gránulos secretores se unen a la membrana durante el proceso de exocitosis y forman parte de la misma se aumenta el volumen de la parte externa de la célula.



El aparato de Golgi se encuentra especialmente desarrollado en células que tienen funciones relacionadas con la secreción de sustancias, como es el caso de las células del sistema nervioso o endocrino.
Como tal, el aparato de Golgi es una de las estructuras que conforman el interior de las células, tanto de organismos animales como de organismos vegetales. Sin embargo, su estructura es más compleja en células animales.


















{kind=link}
{kind=link}